ON
THE PERMIAN/TRIASSIC BOUNDARY IN VIỆT NAM
ĐOÀN NHẬT TRƯỞNG1, ĐẶNG TRẦN HUYÊN1,
NGUYỄN XUÂN KHIỂN1, TẠ HÒA PHƯƠNG2
1Research Institute of Geology and Mineral
Resources, Thanh Xuân, Hà Nội
2Hà Nội University of Natural Sciences, Nguyễn Trãi, Thanh Xuân, Hà
Nội
Abstract: The Meishan D section (Changxing County, Zhejiang, South China) has been ratified as Global Stratotype
Section and Point of the Permian/Triassic boundary. The P/T biostratigraphic
boundary is defined by first appearance of Hindeodus parvus and the
eventostratigraphic boundary - by mass extinction with lithological change. In
Việt Nam, two Permian/Triassic boundary sections have been
preliminary investigated and the conodont Hindeodus parvus has been found at one of them. Based on
the mass extinction of Permian fauna and lithological change recorded from
these sections, the eventostratigraphic boundary at both sections has been
established. These are sections of potential for studying P/T biostratigraphic
boundary in Việt Nam.
I.
INTRODUCTION
The
Permian/Triassic (P/T) boundary is one of chronostratigraphic boundary most
paid attention by geologists, because it is not only the boundary between the
Paleozoic and Mesozoic erathems, but also at this boundary there were great
events of planet scale causing the mass extinction of about 90-95% of genera
and species of the marine and land environments.
Griesbach
(1880) is the first scientist discovering the ammonoid Otoceras in Himalaya and
considered that it belongs to Triassic. Diener (1912) and Spath (1934) are the
first scientists proposing the use of this fossil as marker fauna for
determining the base of Triassic, and thus, the boundary between Paleozoic and
Mesozoic. Later, this propose was accepted and the P/T boundary has been
determined at the base of the Otoceras zone.
According
to Yin Hongfu [6], to the 1984 year the base of the Otoceras zone was still
largely accepted as the base of the Triassic system. However, in some former
years as well as in the 90’ of the past century there still were some authors
having doubts on the age of Otoceras by the following reasons:
1)
There is not evidence on the higher position of the Otoceras zone in
comparing with the Pseudotirolites zone (or Paratirolites zone) – the zone considered as uppermost
Permian, because these zones belong to different paleo-biogeographic provinces.
Therefore, there is not any locality where there is the succession of Pseudotirolites (or Paratirolites) – Otoceras zones
in a same section.
2) The
contact between the Otoceras beds and underlying ones is not continuous in all
places where Otoceras was found.
3) The
lower part of the Otoceras zone bears Permian features, because it contains
clearly Permian ammonoids, conodonts and brachiopods.
Based
on this, since 1986 Chinese geologists proposed to consider the conodont Hindeodus parvus
(Kozur et Pjatakova) as marker fossil for the P/T boundary, in place of Otoceras. The
major part of members of P/T Boundary Working Group (PTBWG) has been supporting
this variant.
The
fossil Hindeodus
parvus has far superiority in comparing with Otoceras. It
is a Tethysian fauna, while Tethys is
the only basin having clearly uppermost beds of Permian. Hindeodus parvus is
easily found in a same section over Pseudotirolites, Paratirolites and
other Tethysian fossils characterizing Permian. It has rather large
distribution, and has been found in China, Kashmir, Salt Range, Transcaucasia, Iran, Austria and Hungary.
Particularly in China, it
has been found in 19 areas of 9 provinces [6].
Figure 1. Location of studied sections
In
1996, 9 members of the PTBWG proposed to select the Meishan section as global
P/T boundary stratotype with the base of Triassic determined by the first
appearance of Hindeodus parvus at the bed 27c. From 1999 to 2000, the
Meishan section passed through 3 tours of selection by the vote of the PTBWG
(Triassic Subcommission, International Commission of Stratigraphy - ICS).
Finally, in March 2001 the Executive Commission of International Union of
Geosciences - IUGS voted the acceptance of Meishan D
section (Changhsing District, Zhejiang
Province) as global P/T boundary stratotype. The P/T boundary is determined at
the base of the bed 27c, where the marker fauna Hindeodus parvus first
appears in the evolutional succession Clarkina changxingensis à Hindeodus latidentatus à Clarkina meishanensis à Hindeodus parvus à Isarciella isarcica, corresponding
to 251 Ma [7].
In the
Meishan section, last fusulinids were extinguished under the H. latidentatus - C.
meishanensis zone (bed 25), while first macrofaunas characterizing
Triassic (ophiceratids, Claraia wangi) do
not occur yet together with Hindeodus parvus, but later, from the bed 28
upward, i.e. over the P/T boundary about 10 cm.
Table1. Zones of conodonts and
ammonoids in the P/T boundary interval
|
|
Conodont zones
|
Ammonoid zones
|
|
TRIASSIC
|
Zone Isarciella isarcica
|
Zone Ophiceras
|
Zone Hindeodus parvus
|
Zone
Otoceras
|
Upper Otoceras bed
|
|
PERMIAN
|
Zone
H. latidentatus – C. meishanensis
|
Lower Otoceras bed
|
|
Zone C. changxingensis yini
Zone C. changxingensis
|
Zone Pseudotirolites - Pleuronodoceras
|
One
can recognized that in the stratotype (as well as in many P/T boundary sections
in South China) the extinction point of time of main Permian fauna groups and
the appearance point of time of Triassic factors do not coincide one with
another. Therefore, there are two boundaries for determination: 1) P/T
boundary, considered as biostratigraphic boundary determined by
the first occurrence of Hindeodus parvus (bed 27c); 2) Eventostratigraphic
boundary determined by the mass extinction of characteristic Permian
fauna groups (including fusulinids) and by the change (usually sudden) of the
lithological composition (bed 25) [3-6]. The biostratigraphic boundary always
lies over the eventostratigraphic boundary by a stratigraphic interval. The
thickness of this interval changes in dependence to the deep- or shallow-water
facies of sediments. In the Meishan and Baoqin sections (Zhejiang Province) this
interval is of 19 and 15 cm in thickness (deep-water facies). In the Taiping
and Heping sections (Guangxi Province) it
is of 90 and 65 cm (shallow-water facies) [3, 5, 7].

Figure 2. Hindeodus parvus (Kozur & Pjatakova) x 200, found at Nhị Tảo section
In
Việt Nam, the
P/T boundary has been first touched upon in the studies of Đặng Trần Huyên [2].
It has been considered as coinciding with the boundary between the Đồng Đăng (P3
dd) and
Hồng Ngài (T1 hn) formations. During the execution process
of the Project “Stratigraphy of Upper Permian - Lower Triassic, forming conditions and
related mineral resources in Bắc Bộ Region” some sections
possible for the study on the P/T boundary have been discovered, among them the
Nhị Tảo and Lũng Pù sections (Fig. 1) are most perspective ones. Especially, in
the Nhị Tảo
section members of the Project have been finding the marker fossil Hindeodus parvus. The
boundary member of the two above sections are characterized by relatively
homogeneous limestone of carbonate shelf facies, therefore, in the field it is
difficult to distinguish the boundary between the two Đồng Đăng and Hồng Ngài
formations. Investigations in these two sections realized at the beginning of
2004 have been allowing to describe in details these sections.
II.
NHỊ TẢO P/T BOUNDARY SECTION
The
Nhị Tảo P/T boundary section is situated near the cistern of the Nhị Tảo Village, in
13 km northwest of the Trà Lĩnh Townlet (Cao Bằng Province). The
section is characterized by carbonate sediments of shelf facies containing in
abundance benthonic faunas as follows (in ascending order):
Bed 1:
organic limestone, dark-grey cherty limestone, 38 cm thick, containing the
foraminiferas Reichelina sp., Nankinella sp., Neoendothyra sp., Dagmarita liantangensis,
Frondina permica, Globivalvulina sp.1, Paraglobivalvulina gracilis.
Bed 2:
dark-grey cherty limestone, 43 cm thick, containing the foraminiferas Dagmarita sp., Frondina sp., Cribrogenerina casta, Cr.
guangxiensis.
Bed 3:
dark-grey organic limestone, 8 cm thick.
Bed 4:
dark-grey cherty limestone, 4.5 cm thick.
Bed 5:
organic limestone, cherty limestone, 41 cm thick, containing the foraminiferas Reichelina sp., Nankinella sp., Baisalina pulchra.
Bed 6:
organic limestone, cherty limestone, 25 cm thick, containing the foraminiferas Reichelina sp., Nankinella sp., Baisalina pulchra.
Bed 7:
dark-grey argillaceous limestone, 10 cm thick, containing the foraminiferas Nankinella pulchra.
Bed 8:
oolitic limestone, 12 cm thick, containing organic residues and the
foraminiferas Reichelina sp., Nankinella sp., Neoendothyra sp., Dagmarita sp., Globivalvulina sp.
1, Baisalina
pulchra, Hemigordius sp..
Bed 9:
dark-grey limestone containing organic residues, 8 cm thick.
Bed 10:
dark-grey limestone, 10.5 cm thick, containing organic residues and the
foraminiferas Reichelina pulchra, Nankinella orbicularia, Dagmarita chanakchiensis, D.
lintangensis, Globivalvulina sp.1, Frondina sp..
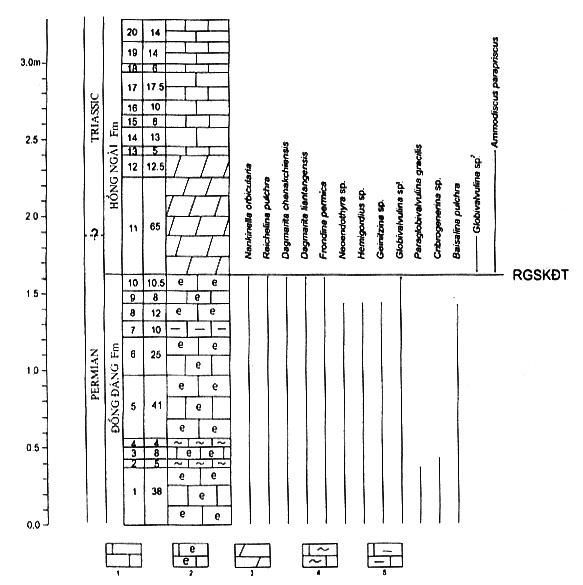
Fig. 3. Stratigraphic column and distribution of foraminiferas
in the P/T boundary section at NhÞ
T¶o
Note: 1. Limestone,2. Organic
limestone, 3. Dolomitic limestone, 4. Cherty limestone,
5. Argillaceous limestone; RGSKĐT:
eventostratigraphic boundary
Bed 11:
grey, irregularly recrystallized dolomitic limestone, 65 cm thick, containing
microgastropods and the foraminiferas Ammodiscus planus, A. parapriscus, Globivalvulina sp.2.
Bed 12:
grey, irregularly recrystallized limestone, 12,5 cm thick, containing
microgastropods.
Bed 13:
grey, poorly dolomitic limestone, 5 cm thick, containing the foraminiferas Globivalvulina sp., Ammodiscus parapriscus.
Bed 14:
grey, poorly dolomitic limestone, 13 cm thick, containing Alga remains.
Bed 15:
grey, poorly dolomitic limestone, 8 cm thick.

Fig. 4. Panorama of the Lũng Pù section
Bed 16:
grey, poorly dolomitic limestone, 10 cm thick.
Bed 17:
grey, poorly dolomitic limestone, 18 cm thick.
Bed 18:
grey, poorly dolomitic limestone, 6 cm thick.
Bed 19:
grey, irregularly recrystallized, poorly dolomitic limestone, 14 cm thick,
containing microgastropods.
Bed 20:
grey, irregularly recrystallized, poorly dolomitic limestone, 14 cm thick.
The
marker fossil Hindeodus parvus (Fig. 2), determined by Tạ Hòa Phương,
was found in a limestone bed lying about 7 m above the bed 20.
For
determining the P/T boundary in a certain area, it needs altogether obey the
standards confirmed in the global stratotype, it means it is obliged to find
conodont fossils in the evolutionary succession Clarkina changxingensis à Hindeodus latidentatus à Clarkina meishanensis à Hindeodus parvus à Isarciella isarcica for
determining the first appearance of Hindeodus parvus. Although in the Nhị Tảo
section the fossil Hindeodus parvus was found, but basing on this only
finding we cannot determine the P/T boundary. Furthermore, in this level Đặng
Trần Huyên [2] has been collecting the bivalve Eumorphotis multiformis,
corresponding to the Claraia stachei zone, which is higher than the Claraia wangi zone.
Therefore, one can say that the bed containing Hindeodus parvus at Nhị Tảo
belongs to the H. parvus zone, but does not correspond to the base of Triassic
system and does not create the standard for determining the P/T boundary.
However,
in the Nhị Tảo
section the eventostratigraphic boundary has been determined on the basis of
the change of lithological and faunal characteristics. They happened between
the beds 10 and 11 (Fig. 3). After the bed 10 all the Permian foraminiferas,
such as Reichelina,
Nankinella, Frondina, Dagmarita, etc., disappear in the section. On the
lithological side, there is a clear change at this level: the section changes
from limestone containing organic residues to dolomitic limestone. This change
reflects an important event in the nature, annihilating in a short time and a
large area many faunal groups, as well as changing the forming conditions of
sediments. On the cause of this event, there has been being different
hypotheses, including even cosmic strike [1], however, this problem will be
touched upon in other occasion.
III. LŨNG PÙ P/T BOUNDARY SECTION
The
Lũng Pù P/T boundary section lies in about 1.5 km west of Sảng Chải Village,
about 7 km southeast of the Mèo Vạc Townlet (Hà Giang Province) (Fig. 4). It is
characterized also by carbonate shelf facies, including following beds (in
ascending order):
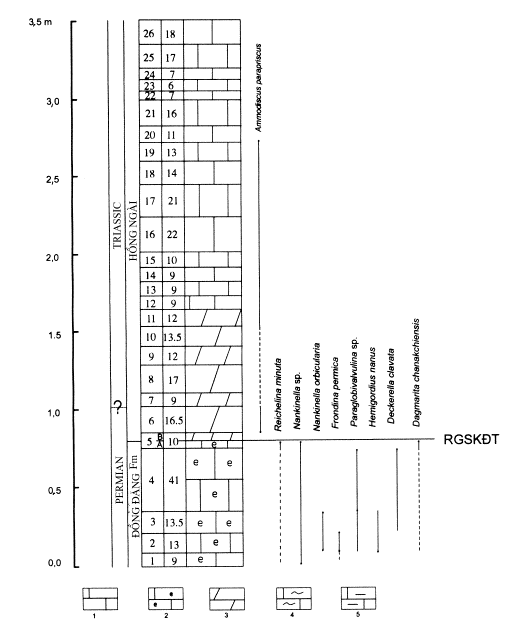
Fig. 5. Stratigraphic column and distribution of foraminiferas
in the P/T boundary section at Lũng Pù
Note: 1. Limestone, 2. Organic
limestone, 3. Dolomitic limestone, 4. Cherty limestone,
5. Argillaceous limestone; RGSKĐT:
eventostratigraphic boundary
Bed 1: grey
limestone bearing organic residues, 9 cm thick, containing the foraminifera Nankinella sp..
Bed 2: grey
limestone bearing organic residues, 13 cm thick, containing the foraminiferas Nankinella orbicularia,
Hemigordius nanus, Frondina permica, Dagmarita sp., Cribrogenerina sp..
Bed 3:
grey limestone bearing organic residues, 41 cm thick, containing the
foraminiferas Nankinella orbicularia, Hemigordius nanus, Paraglobivalvulina sp., Dagmarita sp., Deckerella clavata.
Bed 4: grey
limestone bearing organic residues, 41 cm thick, containing the foraminiferas Paraglobivalvulina sp., Gobivalvulina sp., Pachyphloia sp..
Bed 5a: grey
limestone bearing organic residues, 5 cm thick, containing the foraminiferas Reichelina minuta,
Dagmarita chanakchinensis, Nankinella sp..
Bed 5b:
grey, fine-grained dolomitic limestone, 5 cm thick.
Bed 6:
grey, fine-grained dolomitic limestone, 16.5 cm thick, containing
microgastropods and the foraminifera Ammodiscus parapriscus.
Bed 7:
grey, irregular-crystalline dolomitic limestone, 9 cm thick.
Bed 8:
grey, rich in dolomite limestone, 17 cm thick.
Bed 9: grey
dolomitic limestone, 12cm thick.
Bed 10: grey
dolomitic limestone, 13.5 cm thick.
Bed 11:
grey, rich in dolomite limestone, 12 cm thick, containing microgastropods and
the foraminifera Ammodiscus parapriscus.
Bed 12: grey
dolomitic limestone, 9 cm thick, containing microgastropods and the
foraminifera Ammodiscus parapriscus.
Bed 13: grey
limestone, 9 cm thick.
Bed 14:
grey, partially dolomitized limestone bearing organic residues, 9 cm thick,
containing the foraminifera Ammodiscus parapriscus.
Bed 15:
grey, fine-grained dolomitic limestone, 10 cm thick, containing microgastropods
and the foraminifera Ammodiscus parapriscus.
Bed 16:
grey, poor in dolomite limestone, 22 cm thick, containing microgastropods.
Bed 17:
grey, irregular-crystalline dolomitic limestone, 21 cm thick, containing alga
and microgastropod remains.
Bed 18:
grey, poor in dolomite limestone, 14 cm thick, containing alga and
microgastropod remains.
Bed 19:
grey, poor in dolomite limestone, 13 cm thick, containing microgastropods and
the foraminifera Ammodiscus parapriscus.
Bed 20:
grey, poor in dolomite limestone, 27 cm thick.
Bed 21:
grey, poor in dolomite limestone, 7 cm thick, containing microgastropods.
Bed 22:
grey, poor in dolomite limestone, 6 cm thick.
Bed 23:
grey, poor in dolomite limestone, 42 cm thick, containing microgastropods.
In the
Lũng Pù section, the P/T biostratigraphic boundary has not been also
determined, because conodont fossils have not been found yet, and only the
eventostratigraphic boundary has been determined. It lies between the 5a and 5b
bed (Fig. 5). The beds from 1 to 5a contain in abundance Permian foraminiferas,
such as Reichelina,
Dagmarita, etc.. They disappear from the bed 5b upward. On the
lithological side there is also a change between the two 5a and 5b beds. The
beds from 1 to 5a consist mainly of limestone bearing organic residues, while
from the bed 5b upward there is only fine-grained dolomitic limestone; the
contact surface between the two rock types is of suture form (Fig. 6). As in
the Nhị Tảo
section, the change between the two 5a and 5b beds in the Lũng Pù section is
the imprint of an important event in the nature, causing the mass extinction of
many organic groups and the change of forming conditions of sediments.

Fig. 6. Eventostratigraphic boundary
(black line) in the lamella from Lũng Pù
IV.
DISCUSSION AND CONCLUSIONS
The
two above described sections show that only the P/T eventostratigraphic
boundary has been determined. It is also the boundary between the two Đồng Đăng
and Hồng Ngài Formations. The posed problem is that, in Việt Nam (concretely,
the two Nhị Tảo and
Lũng Pù sections) is there the P/T biostratigraphic boundary? There are two
possibilities:
1)
There is a stratigraphic gap at the eventostratigraphic boundary. In this case
there is only this boundary, as in the case of the Hồng Ngài section. There,
beds of argillaceous limestone
containing Claraia wangi, Glyptophiceras sp. belonging to the base of the
Hồng Ngài Formation rests directly upon limestone containing Late Permian
fusulinids of the Đồng Đăng Formation;
2)
There was a continuous sedimentary deposition or inconsiderable gap at the
eventostratigraphic boundary. In this case, over it there will be the
biostratigraphic boundary. This lies in the basal part of the Hồng Ngài
Formation, that means the P/T boundary does not coincide with that between the
two Đồng Đăng and Hồng Ngài Formations.
Correlating
with the Taiping P/T boundary section, Guangxi, China [3] situated in about 150
km northeast of Nhị Tảo in the same limestone bands extending from China to
Việt Nam we notice that the Nhị Tảo and Lũng Pù sections have the same type of
section with the same lithological and faunal composition. There is much
possibility that the two Nhị Tảo and
Lũng Pù sections belong to the above said second case, it means the P/T
biostratigraphic boundary can be found over the eventostratigraphic boundary a
certain interval in the section. The continuation of investment for more
detailed study on this possibility is very necessary.
Acknowledgement.
The paper is realized with the support of the Basic Research
Program and the contribution of precious ideas from colleagues of the Division
of Paleontology and Stratigraphy, Research Institute of Geology and Mineral
Resources. The authors express their deep thanks.
REFERENCES
1. Benton
M.J., and R.J. Twitchett, 2003. How to kill (almost) all life: The end-Permi an
extinction event. Trends in Ecology and Evolution, 18/7.
2. Đặng Trần Huyên, 1998.
Lower Triassic bivalves from Hồng Ngài
Formation (Sông Hiến structural zone). J. Geology, B/11-12 : 95-106. Hà Nội.
3. Lehrmann D. J., J.L. Payne, S.V.
Felix, P. M. Dillett, Hongmei Wang, Youyi Yu, and Jiayong Wei, 2003. Permian-Triassic boundary sections from
shallow-marine carbonate platforms of the Nanpanjiang Basin, south China: Implications for oceanic conditions associated with the end-Permian
extinction and its aftermath. Paleobios, 18: 138-152.
4. Metcalfe Ian, R.S. Nicoll, R. Mundil, C. Foster, J. Glen, J. Lyons,
Wang Xiaofeng, Wang Cheng Yuan, P. R. Renne, L. Black, Qu Xun and Mao Xiaodong,
2001. The Permian-Triassic boundary
and mass extinction in China. Episodes, 24/4 : 239-244.
5. Wang Cheng-Yuan, 1994. A conodont-based high-resolution eventostratigraphy
and biostratigraphy for the Permian-Triassic boundary in South China. In Permian
stratigraphy, environment and resources. Vol. 1, Paleontology &
Stratigraphy. Nanjing University Press.
6. Yin Hongfu, 1994.
Reassessment of the index fossils at the Paleozoic-Mesozoic boundary. In Permian stratigraphy, environment and
resources. Vol. 1, Paleontology & Stratigraphy. Nanjing University Press.
7. Yin Hongfu, Zhang Kexin, Tong
Jinnan, Yang Zunyi and Wu Shunbao, 2001. The global stratotype section and point (GSSP) of
the Permian-Triassic boundary. Episodes, 24/2.